


It’s hard to pin down when the idea dawned. Apparently, it was already common among Canadians in 1912 when Ernest Thompson Seton considered the phenomenon of population swings among snowshoe hares:
“The species increases far beyond the powers of predaceous birds or beasts to check,” he wrote in Arctic Prairies, “and the Rabbits after 7 or 8 years of this are multiplied into untold millions. . . . Finally they are so extraordinarily superabundant that they threaten their own food supply as well as poison all the ground. A new influence appears on the scene, and the Rabbits are wiped out. . . . [W]hen the destruction is complete it is an awful thing for the carnivores that have lived on the bunny millions and multiplied in ratio with their abundance.”
Opinion
“[T]he Canada Lynx lives on Rabbits,” he continued, “increases with them, and on their failure dies of starvation in the unrabbited woods.”
Seton may have been the first to consult the records of the Hudson’s Bay Company to find out how the trade in lynx pelts changed over time. In 1900, he reported that the company took in 4,473 lynx skins. In 1906, it bought 61,388. In 1908, the take dropped to 9,664 — the ten-year cycle of snowshoe hares and lynx, which, at that time, was reflected in more than a century of the business records of the great North American fur monopoly.
Humans have recognized the ebb and flow in populations of wild animals for as long as they’ve hunted, but this predictable rise and fall in numbers of snowshoe hares was something different, clearly nonrandom and endlessly fascinating for the emerging generation of zoologists.
In 1924, the young Oxford don Charles Elton gathered information from around the world and broadened the consideration of cycles in wildlife populations to include several species of lemmings and their predators, snowshoes hares and their predators, jackrabbits, several species of mice, muskrats, marmots, fishers, mink, wolverine, skunks, sandgrouse, waxwings, redpolls, crossbills, nutcrackers and a variety of insects.
By the mid-1940s, some writers in the field of wildlife ecology had further expanded the list of species whose populations cycle. In his 1949 book, The Way to Game Abundance: An Explanation of Game Cycles, Wisconsin naturalist Wallace Grange assumed that there was one cycle driving changes in a vast array of wild animals. He called it “the game cycle,” and included all rabbits and hares, squirrels, foxes, wolves, badgers, skunks, mink, great horned owls, all northern species of grouse, mice, lemmings, ground squirrels, pheasants, Hungarian partridge and bobwhite quail.
According to Grange, “cycles are so common that it is perhaps easier to name events which are not cyclic than to run through a whole list of natural events, most of which are cyclic. The dictionary defines cycle as: ‘An interval of time occupied by one round of events recurring in the same order in a series.’ This definition does not depend upon regularity, or upon a constant interval length. It depends upon recurrence in the same order. . . . A cycle can show intervals of 3,4, 5 or other differing numbers of years and still be a ‘regular’ cycle.”
So, according to Grange, a cycle in wildlife populations didn’t have to occur in a set period of time and, by implication, didn’t have to return to a starting point, two characteristics the dictionary definition he quotes seems to require.
Lamont Cole, a zoology professor at Cornell University, was skeptical of this all-encompassing view. Five years after Grange published his discussion of cycles in wildlife, Cole wrote: “Since the term ‘cycle’ has come to signify (at least among biologists and economists) a sequence of events repeating in a definite order but without any necessary implication of strict regularity in time, and since populations obviously cannot continue to either increase or decrease indefinitely, it seems that when we speak of a ‘population cycle’ we are stating little or nothing beyond the fact that the population does not always remain exactly at a constant state.”
In his 1963 book, Wildlife’s Ten-Year Cycle, Lloyd Keith, a wildlife ecology professor at the University of Wisconsin, spent several pages grappling with the definition of cycle as it applied to changing populations of wildlife. What was a “regular interval” in a population cycle? Keith wondered. And how could an analyst tell the difference between a relatively minor change in the size of a population and a true peak or valley? How many times does a population have to rise and fall before an observer can call it a legitimate cycle and not just a random fluctuation in numbers? Keith concluded that three- to five-year changes in numbers, like those of lemmings in the Arctic, were as likely to be random fluctuations as cycles. The nine- to 11-year cycle of numbers among snowshoe hares and their predators, as well as a similar cycle in many ruffed grouse populations, were decidedly not random, he concluded.
He spent the next 23 years with a succession of graduate students investigating the mechanisms that drive the snowshoe hare cycle in Canada.

The phenomenon of cycles in wildlife populations has generated lively discussion in the technical community: Do cycles exist? If they do, are they more than random fluctuations? How do we recognize them? And what might be their cause? Ecologists and population biologists find the search for answers to these questions endlessly absorbing. For most people, the possibility of cycles in the numbers of wild animals is, at most, an oddity of nature — a footnote in a BBC documentary, mildly entertaining, quickly forgotten.
But the debate takes on far more force when it involves a species in decline. A species that has been petitioned for listing as “threatened” under the provisions of the federal Endangered Species Act. A species like the sage grouse.
Over much of the first half of the twentieth century, conservationists were less concerned about cycles in sage grouse populations than whether any populations could survive. In 1932, the ornithologist Arthur Cleveland Bent had this to say about the trajectory of sage grouse numbers:
“It is disappearing very fast, notably in California, Oregon, and Washington. . . . To save this fine bird from extinction, as civilization spreads, the open season for shooting it must be shortened and the bag limits reduced. Even then, it probably can not be saved except on protected reservations.” Most states in sage-grouse range followed Bent’s advice and closed sage-grouse seasons for several years in the 1930s and 1940s.
Probably the first observer to advance the idea of sage grouse cycles was Wallace Grange, the Wisconsin naturalist, who was apparently anxious to flesh out his list of game animals involved in his theory of “the game cycle.” Chances are good that he had never seen a sage grouse, but he assumed that this grouse would follow the same pattern he thought he saw in every other North American game bird.
He was apparently unaware of George Girard’s pioneering study of sage grouse, published in 1937, which never mentioned the possibility of a cycle in sage grouse numbers, and with good reason, since sage grouse had been in catastrophic decline for 40 years when Girard began his research. In fact, Girard’s research was driven by concern over that decline.
But, by 1952 when Robert Patterson’s landmark monograph, The Sage Grouse in Wyoming, was published, the idea of cycles had found its way west: “It would seem highly probable,” Patterson wrote, “that this species of grouse may also be exhibiting the cyclic phenomena or oscillations so well associated with other grouse and upland game birds.” Patterson had no wide-scale, long-term data to support this assertion. He based his conclusion on 10 years of live-trapping records in Wyoming’s Eden Valley, on seat-of-the jeans reports from other sage-grouse states, and, in all likelihood, on Grange’s confident assertions.

As the years have gone by, as wildlife research has become more sophisticated, as techniques for collecting population data have advanced, the list of species involved in Grange’s “game cycle” has been whittled down. There’s solid research to support the validity of the venerable snowshoe hare cycle in Canada, and comparable support for the existence of a ruffed-grouse cycle in the Great Lakes states (although not in peripheral populations in the West or Appalachians). But these days, there’s no reason to believe that populations of prairie chickens, sharp-tailed grouse, dusky grouse, or sooty grouse go through cycles. Or pheasants. Or bobwhite quail. Or gray partridge. Or foxes. Or jackrabbits, wolves, coyotes, snowy owls, or marsh hawks. Grange thought all these species were part of the “game cycle.” Further investigation showed they are not.
Over the last 70 years, the notion that sage-grouse populations have a 10-year cycle has been accepted with remarkably little technical rigor. Sage-grouse populations are estimated by counting the number of males on communal breeding grounds each spring and comparing those numbers through time. There are significant problems with using these counts as real population estimates, but they’re the best we’ve got, so they’re used with the understanding that they are only a rough approximation of reality.
The best counts available reach back a little more than 50 years or about five of the proposed 10-year cycles, which is far less than the data that have been amassed on ruffed grouse or the classic information on Canadian snowshoe hares from the records of the Hudson’s Bay Company, which now stretch back 150 years. The “peaks” in sage-grouse counts that supposedly indicate a cyclic high can be anemic, so anemic, in fact, as to raise the question of whether they’re real peaks or just blips in the data, and the counts are often “smoothed” statistically to get rid of inconvenient year-to-year changes that might not be consistent with the expected pattern of the cycle.
Three of the most recent attempts to analyze sage-grouse populations reach startlingly different conclusions. Jeffrey Row and Bradley Fedy with the University of Waterloo in Ontario looked at 50 years of spring counts and, after smoothing the data, concluded that “most populations (11 of 15) were cyclic at some point in a 50-year time series (1965-2015), but the patterns varied over both time and space. Several populations demonstrated amplitude dampening or loss of cyclicity following population lows in the mid-1990s. Populations in the core of the range had more consistent cyclic dynamics, but period length appeared to shorten from 10-12 to 6-8 years.”
So, the data covered just five or six possible peaks in the proposed cycle. In that limited span, 11 of the 15 populations showed some sign of a cycle, but, of those, some didn’t show cycles after the mid-1990s, and others showed a change of four years — a 50% to 60% difference — in the time span of the supposed cycle. And those patterns, such as they were, emerged after the original data had been “smoothed.”
A markedly different picture emerges from another analysis, this one by Edward O. Garton, professor of statistics at the University of Idaho, and several colleagues. In 2011, they used the spring breeding data from 1965 through 2007 for each discrete population to estimate and graph the minimum number of males in each. The resulting graphs show no perceptible sign of a cycle. A few of the graphs show stable populations at very low numbers; most show steady decline. The analysis appears in “Greater Sage-Grouse Population Dynamics and Probability of Persistence” in the book, Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats.
In a parallel report, Garton concluded that “Our results combined with findings from other recent studies suggest sage-grouse populations that are quite small or exposed to continuing severe threats (wildfire, energy development) . . . continue to decline in spite of various conservation efforts. Populations occupying landscapes where wildfire is relatively rare and energy development limited have fared better . . . but nowhere have we found evidence that any larger populations are stable to increasing. Conservation efforts that emphasize protecting remaining habitat over broad landscapes are necessary to insure sage-grouse persistence on these lands.”
The most recent analysis of sage-grouse population trends, undertaken by the U.S. Geological Survey and published in 2020, supports Garton’s conclusions. The authors of the study reported “37.0, 65.2, and 80.7-percent declines in abundance range-wide during short (17 years), medium (33 years), and long (53 years) temporal scales, respectively.” They found evidence of increasing trends in some areas over recent decades, but their models predicted that more than three-quarters of spring breeding leks had a 50-50 chance of disappearing over the next half-century.
That’s what the statistics tell us.
This would all be utterly academic if it weren’t for the way the idea of a sage-grouse population “cycle” affects management and conservation for the bird. When the spring counts start to go down, the reaction from politicians with a special interest, along with a startling number of wildlife professionals, is, “That’s just the cycle.” If the “cycle” is supposed to last 10 to 12 years, that explanation is good for at least six years, maybe a bit longer.
And, when the “cycle” hits bottom, there’s the expectation that the population will bounce back. The tendency is to look at the highs in the population graph and discount the lows, which may not make much difference if the wildlife species in question numbers in the tens of millions, but it’s dangerous in the extreme if the species — like sage grouse — has suffered major losses and continues to decline. If the species ever disappears, it will be at a low point in the graph, not a high — that’s where our focus should be. As Garton’s analysis suggests, if the current trajectory of greater sage-grouse populations across the West continues unabated over the next century, we’ll be left with less than 5% of the birds we have now. Functional extinction.
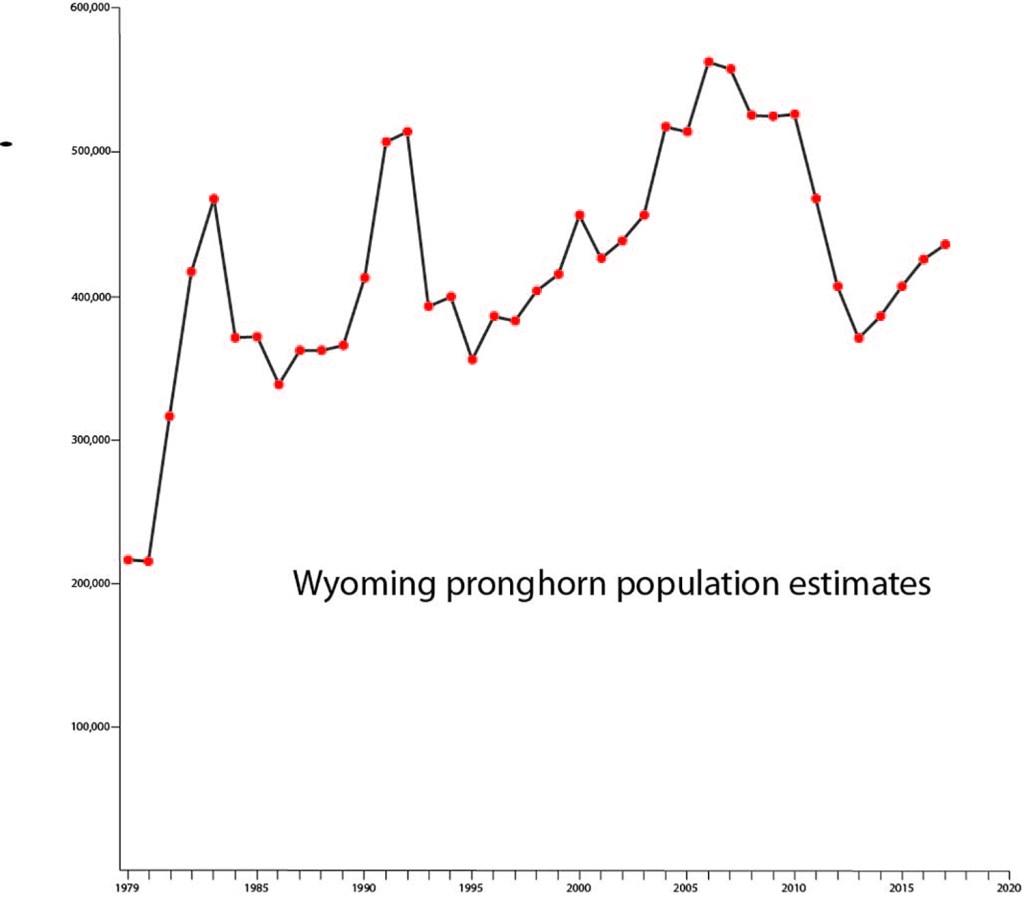
It’s interesting that population estimates of mule deer and pronghorn in Wyoming over the last 40 years show distinct peaks at roughly 10-year intervals, yet, when it comes to habitat management or hunting license quotas, no one talks in terms of 10-year cycles. The lows generally attract more attention than the highs, and, in the end, the conversation among wildlife professionals and concerned citizens is all about improving habitat to support more animals, which is as it should be. Why should our approach to sage grouse conservation be any different?
The thing that makes a cycle a cycle, the one characteristic that sets it apart from other kinds of change, is that a series of events leads eventually back to a starting point. Sage grouse numbers haven’t gotten back to their starting point in well over 100 years. The concept of the cycle does nothing to help us solve the fundamental problems facing sage grouse in the West. It’s time we dropped the comforting myth and dealt, clear-eyed, with the troubling reality of a trend that shows no lasting sign of improvement.
Thank you for a well-researched and thoughtful article. What can we do to be helpful?
I am happy to see this important topic getting some attention in the media. However, much of Mr. Madson’s description of sage-grouse population trends is a mischaracterisation and misunderstanding of the current literature. As a research scientist, I have led and collaborated on much of the peer-reviewed literature addressing the issue of population cycles in sage-grouse, both in Wyoming and range-wide. I have submitted a detailed response to Mr. Madson’s claims to the editorial office at Wyofile and hope they agree to publish it in the near future.
Dr. Brad Fedy
Associate Professor
bfedy@uwaterloo.ca
Awesome article. There are some additional major impacts to wildlife populations which are mostly known to Wyoming ranchers – the folks who live on the land. They include periodic KILLER BLIZZARDS which absolutely decimate wildlife populations and occur approximately every 8-15 years. Next is el Nino weather patterns which cause field mice populations to explode and thence hanta virus human deaths on the Navajo reservation – quite cyclic in nature. And last but not least, is the introduction of sylvatic plague into our wildlife populations in the early 1950s. This new influence on the cyclic nature of wildlife populations is mostly known to ranchers who have observed it first hand. I witnessed a complete cycle of wildlife population fluctuations in the 1990s in Niobrara County and only say that it is not understood by the academic community and Game and Fish. You have to live out there to properly understand the affect of sylvatic plague.
When Game and Fish surveys wildlife populations, the survey should always note whether or not that the population numbers were observed after a KILLER WINTER, after an outbreak of sylvatic plague in the area or whether there had been a wet/ el Nino year recently; and in some cases, several of these events occur within a 1-2 year period.
Yes, wildlife populations are cyclic, but understanding the under lying causes is best left to the ranchers – they know.
In watching wildlife numbers, birds, deer, pronghorn etc. Environmental conditions seem to impact animals livelihood to a far greater degree than we understand. Drought during chick rearing, no snow, abnormal low temperatures during winter.
A govt trapper told me when he trapped the bobcats out of sheep rancher range the sage hen populations crashed.
I found your article very informative.
Thankyou
Chris, you have produced a paper that should be a mandatory read for every aspiring wildlife student on the planet earth. In my life time “Cycles ” are a get out of jail free card for every politician and in some cases unfortunately, wildlife managers.Great piece of work Chris.
Chris Madison is a helluva asset to our state, a true Wyoming treasure. Clear thinking, a high elevation perspective, and apolitical writing yields the best summary on the Sage grouse/habitat issue I have ever read.
I only wish that he had been Director of the Game and Fish. Wyoming’s wildlife populations would be in much better shape.
Reading over & over how “energy development” (drilling & mining?) and grazing of livestock doesnt affect these birds is laughable. This is an interesting article and sure does seem to make the idea that their low numbers have nothing to do with “cycles”. Thats just been an excuse.
Thanks for addressing the issues around “cycles” in wildlife. Daniel B. Botkin wrote about this and other assumptions in the field of ecology in his two books “Discordant Harmonies” and the updated version ” The Moon in the Nautilus Shell.” He actually specifically talks about the snowshoe hare/lynx cycle and the data from the Hudson Bay Company. There are a lot of foundational myths and scientific concepts taken out of context that still appear in policy making, common understandings, and even the field of ecology.
At every scale of measurement, sage-grouse have been in decline since settlers of European descent moved into their occupied range and began altering it. Even relatively “intact” habitat has been altered sufficiently to tip the balance away from cryptic shrubsteppe obligates like sage-grouse, in favor of their predators – winged, and 2 and 4 legged mammals. Sage-grouse are one of the few species where the road hunting is as good the last day of the season as it was the first. Even if rangewide harvest moratoriums were implemented and no more habitat converted or degraded, I fear fenceposts and 2-track roads may be sufficient to drive the species to extinction in the next 100 years.
Great article! Lots of information supported by excellent references. Note how
“cycle” concept is also used by those who reject climate change science.
Great observation
Chris continues to challenge the political, multiple use concepts of management. This will ruffle some feathers.
We have been lulled with language like “it’s just in the down part if a cycle.”
Like blue grouse that used to be extremely abundant and now aren’t. We’ve been in the low cycle for too long. Yet ,seasons rarely change.
Game and Fish should not be part of Governor’s cabinet. The director should be hired by the board and the agency as independent as possible. Let them be advocates for wildlife independent of politics.
Politicians like to frame wildlife issues in palatable ways. But, hard choices have to be made. With more and more demands on public lands, we can not multiple use all of it.